Monday, November 06, 2006
「癢」的神經生物機制(Neurobiology of itch)!
在坊間的教科書中,「癢」的神經機制是較少數被科學家探究的領域範疇。其原因主要是因為沒有恰當的動物模式(animal model)可以供實驗者做系統性的操弄與觀察,動物不會主動告知實驗者「我會癢!」這件事,取而代之的是抓(scratch)或摩擦的動作反應,這些反應不易觀察且具爭議性;隨這現代神經科學技術的發展,如活體電生理(in vivo electro-physiological recording)紀錄、大腦功能造影(functional brain imaging)等技術的使用和進步,這些限制正逐漸地被克服。「癢」和「痛」有類似的的神經受體引發嗎?還是個別有特定的神經迴路負責?以下的整理將提供讀者對這感官「癢」有初步的認識。

(一)皮膚發炎疾病引發的癢:如蚊子咬、皮膚過乾、濕疹或牛皮癬等皮膚病所引發的「癢」。
(二)系統性疾病引發的癢:如甲狀腺抗進或長期肝、腎功能不好的患者會有「癢」的併發症。
(三)神經性的癢:如骨刺壓迫末梢神經或截肢手術後病人常報告的「癢」。
(四)心因性的癢:如精神病患的寄生蟲幻覺、長期壓力引發或憂鬱症病人所抱怨的「癢」。
從這裡我們也可以看出「癢」的多源性,從周邊(PNS)到中樞神經系統(CNS)都可以引發「癢」的感覺,加上「癢」也有急性(acute)和長期(chronic)複雜度的不同。這些特性和現階段科學家對「痛覺」的了解有若干的相似。所以近代神經科學對「癢」的研究報告幾乎都來至於傳統做「痛覺」的實驗室。
(二)發炎組織附近的免疫細胞會分泌蛋白酶( protease),過多的蛋白酶會活化神經末梢上的PARs(protease-activated receptors),目前已知的PARs有多種,其中已經證實PAR2和「皮膚癢」的引發有關係。首先,在有「皮膚癢」報告的皮膚炎病患身上,發炎部位的PAR2表現量會升高;其次局部活化皮下PAR2會讓受試者報告「癢覺」。
(三)在周邊神經末梢分布大量的TRP(transient receptor potential)受體,這些受體本身就是離子通道(ion channel),活後直接影響末梢神經的神經電位,在痛覺研究中已經被證實和冰或熱所引發溫度痛覺(thermal pain)有密切關係,TRP受體有很多種,其中TRPM8(一種cold receptor)被認為和冰敷的止癢效果有關係;TRPV1和TRPV3和樟腦局部塗抹後,溫熱且止癢的效果有關係。
(四)嗎啡與大麻受體也和「癢覺」有關係,內生性腦啡(opioids)和大麻素(cannabi-noids)作用在其受體(μ-,κ-,δ- CB1 or CB2)後不只有止痛,也有止癢的效果。惟活化受體的層次不同(skin、spinal或 supra-spinal level),其「止癢」和「使癢」的效果迥異。舉例來說,在脊椎活化μ-受體會產生癢的感覺,而活化κ-受體有止癢的報告。
(一)這些傳遞癢覺的細胞沒有自發性(spontaneous)電位活動。
(二)這些細胞之所以沒有電位活動懷疑是被傳遞痛覺的細胞活動所抑制( pain-induced inhibition)。
(三)這些細胞從脊髓的lamina I 直接投射到ventrocaudal part of the nucleus medialis dorsalis( MDvc),MDvc在向前投射到前扣帶回皮質(anterior cingulate cortex)與背側島狀皮質(dorsal insular cortex)。
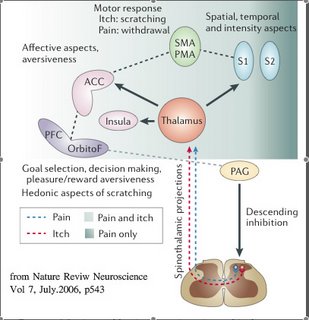
在1990年代初期的一系列人腦造影研究中,我們已經知道痛覺在中樞皮質的處理分區有其時間、空間、情緒、動機、強度等....許多不同的向度。那關於「癢覺」呢?台北榮總的謝仁俊醫師可是全世界第一個(1994)用PET來回答這問題的科學家,他發現前額葉皮質(prefrontal cortex)、前運動皮質(pre-motor area)和前扣帶回皮質(anterior cingulate cortex)對「癢覺」有反應。後來的fMRI也有類似的發現。和「痛覺」比較,這些對「癢覺」有反應的中樞皮質幾乎是重複的;換句話說,到目前為止,大腦中並沒有哪各區域被證實對「癢覺」的處理有專一性!
在坊間的教科書中,「癢」的神經機制是較少數被科學家探究的領域範疇。其原因主要是因為沒有恰當的動物模式(animal model)可以供實驗者做系統性的操弄與觀察,動物不會主動告知實驗者「我會癢!」這件事,取而代之的是抓(scratch)或摩擦的動作反應,這些反應不易觀察且具爭議性;隨這現代神經科學技術的發展,如活體電生理(in vivo electro-physiological recording)紀錄、大腦功能造影(functional brain imaging)等技術的使用和進步,這些限制正逐漸地被克服。「癢」和「痛」有類似的的神經受體引發嗎?還是個別有特定的神經迴路負責?以下的整理將提供讀者對這感官「癢」有初步的認識。
- 「癢」的分類

(一)皮膚發炎疾病引發的癢:如蚊子咬、皮膚過乾、濕疹或牛皮癬等皮膚病所引發的「癢」。
(二)系統性疾病引發的癢:如甲狀腺抗進或長期肝、腎功能不好的患者會有「癢」的併發症。
(三)神經性的癢:如骨刺壓迫末梢神經或截肢手術後病人常報告的「癢」。
(四)心因性的癢:如精神病患的寄生蟲幻覺、長期壓力引發或憂鬱症病人所抱怨的「癢」。
從這裡我們也可以看出「癢」的多源性,從周邊(PNS)到中樞神經系統(CNS)都可以引發「癢」的感覺,加上「癢」也有急性(acute)和長期(chronic)複雜度的不同。這些特性和現階段科學家對「痛覺」的了解有若干的相似。所以近代神經科學對「癢」的研究報告幾乎都來至於傳統做「痛覺」的實驗室。
- 關於「癢」的理論
- 周邊神經系統的「癢覺」
(二)發炎組織附近的免疫細胞會分泌蛋白酶( protease),過多的蛋白酶會活化神經末梢上的PARs(protease-activated receptors),目前已知的PARs有多種,其中已經證實PAR2和「皮膚癢」的引發有關係。首先,在有「皮膚癢」報告的皮膚炎病患身上,發炎部位的PAR2表現量會升高;其次局部活化皮下PAR2會讓受試者報告「癢覺」。
(三)在周邊神經末梢分布大量的TRP(transient receptor potential)受體,這些受體本身就是離子通道(ion channel),活後直接影響末梢神經的神經電位,在痛覺研究中已經被證實和冰或熱所引發溫度痛覺(thermal pain)有密切關係,TRP受體有很多種,其中TRPM8(一種cold receptor)被認為和冰敷的止癢效果有關係;TRPV1和TRPV3和樟腦局部塗抹後,溫熱且止癢的效果有關係。
(四)嗎啡與大麻受體也和「癢覺」有關係,內生性腦啡(opioids)和大麻素(cannabi-noids)作用在其受體(μ-,κ-,δ- CB1 or CB2)後不只有止痛,也有止癢的效果。惟活化受體的層次不同(skin、spinal或 supra-spinal level),其「止癢」和「使癢」的效果迥異。舉例來說,在脊椎活化μ-受體會產生癢的感覺,而活化κ-受體有止癢的報告。
- 傳遞「癢覺」的神經迴路(貓的神經解剖實驗)
(一)這些傳遞癢覺的細胞沒有自發性(spontaneous)電位活動。
(二)這些細胞之所以沒有電位活動懷疑是被傳遞痛覺的細胞活動所抑制( pain-induced inhibition)。
(三)這些細胞從脊髓的lamina I 直接投射到ventrocaudal part of the nucleus medialis dorsalis( MDvc),MDvc在向前投射到前扣帶回皮質(anterior cingulate cortex)與背側島狀皮質(dorsal insular cortex)。
- 傳遞「癢覺」的中樞迴路(人的腦部造影實驗)

在1990年代初期的一系列人腦造影研究中,我們已經知道痛覺在中樞皮質的處理分區有其時間、空間、情緒、動機、強度等....許多不同的向度。那關於「癢覺」呢?台北榮總的謝仁俊醫師可是全世界第一個(1994)用PET來回答這問題的科學家,他發現前額葉皮質(prefrontal cortex)、前運動皮質(pre-motor area)和前扣帶回皮質(anterior cingulate cortex)對「癢覺」有反應。後來的fMRI也有類似的發現。和「痛覺」比較,這些對「癢覺」有反應的中樞皮質幾乎是重複的;換句話說,到目前為止,大腦中並沒有哪各區域被證實對「癢覺」的處理有專一性!
# posted by shinghonglin @ 7:38 AM 
![]()